
Picture 1 Nepenthes bicalcarata
“The pitchers of Nepenthes possesses extraordinary power of digestion” … “The fact that a plant should secrete, when properly excited, a fluid containing an acid and ferment, closely analogous to the digestive fluid of an animal, was certainly a remarkable discovery.” (Charles Darwin, Insectivorous Plants pg. 97).

Nepenthaceae Dum. famous within the carnivorous kingdom for its characteristic insect catching pitchers, is thought to only contain one genus Nepenthes L. Although some have split one species (Nepenthes pervillei) into another genus Aneurosperma due to its distinct seed and floral morphology (Heywood et al. 2007). However, this taxonomy is rarely followed. Many Nepenthes species circa 87 sp are found in Borneo or Sumatra with the rest of the species having a Palaeotropical distribution (Clarke 1997). There are an estimated 140species currently known.
The species within the family typically thrive in low nutrient conditions such as montane, disturbed forests and sometimes in grasslands. They can tolerate altitudes from sea level up to 2500m (Cheek & Jebb. 2001). The climbers amongst the Nepenthes use tendrils to support and anchor themselves to the surrounding vegetation. If they are not climbers they can be epiphytic. The key diagnostic features for this family of perennials are: a specialised leaf midrib developing into tendrils followed by pitchers and a terminal inflorescence of unisexual four-tepalled flowers arranged in cymes (Cheek & Jebb. 2001).
A little more morphology
Typically the Nepenthes have different growth stages which then determines the stem height. The rosette stage is where most of the Nepenthes begin, producing short stems (<2m) as the Nepenthes matures, it may go to the liana phase where the stem can grow as long as 20m, these mature stems give rise to different lower and upper pitchers (Heywood et al. 2007). However, not all Nepenthes species complete these stages for example N.argentii flowers within the rosette phase whereas N. raja does not have climbing stems but instead flowers from the shorter stems (Cheek & Jebb 2001). The leaves have no stipules and are often alternate petiolate or can be sessile (attached without a petiole).
Nepenthes are dioecious, containing male and female inflorescences on separate plants, often arranged in cymes although these can be in a panicle (Cheek & Jebb 2001). The flowers themselves are small red/yellow/green, 4-merous and generally bracteate or ebracteate (without bracts), with male flowers containing many stamens (4-24). The female tepals are typically nectar secreting (Heywood et al. 2007).
For the majority of Nepenthes species the leaf midrib extends into a tendrils which will develop into a pitcher, where size and colouration will very much depending on the species (picture 3). These highly sophisticated pitfall traps primary aim is to catch prey, using a variety of mechanisms. The typical prey found in the pitcher consist of insects; ants being the most popular in a majority of Nepenthes species. However, in N. bicalcarata, ants apparently have a mutualistic relationship with the Nepenthes species.

Picture 2: Bat seen roosting in N. stenophylla pitchers. Attributes to Alastair Culham 2013
It was found that a particular ant species lived in the hollowed out tendrils of N.bicalcarata, living on the surrounding insects while helping to break down the larger prey (Clark. 1997). Other animals have also been sighted to use the pitchers such as Monkeys, quenching their thirst from rainwater in the lowland species N. rafflesiana (Clark. 1997). Moreover, bats have been recently spotted nesting in the empty pitches of Nepenthes rafflesiana and now in Nepenthes stenophylla (picture 2). Surprisingly, it appears an unusual mutualistic relationship between the bats and the pictures has developed, this is rather out of character for Nepenthes species, where the main aim is to catch prey rather than allow cohabitation and shelter ( http://rsbl.royalsocietypublishing.org/content/7/3/436.short).
Once the pitchers have reached full maturity the previously sealed lid will finally open, unveiling the pitfall trap below. Firstly, the peristome (pitcher lip) will have sweet nectar coating the surface, dispensed from the nectar glands located on the underside of the lid, in order to entice the prey (Picture 3). The outer structure to the pitcher usually contains a number of chlorophyll cells which oxygenate the fluid, preventing the digestive juices from fermenting and the wall is lined with a copious supply of veins (Cheek & Jebb 2001). Just below the peristome lies a waxy coating that reduces the insects ability to grasp onto the pitcher. Ultimately this will result in the insect falling into the digestive fluids composed of enzymes which will break down the insect and enable the uptake of nutrients (Clarke 1997).
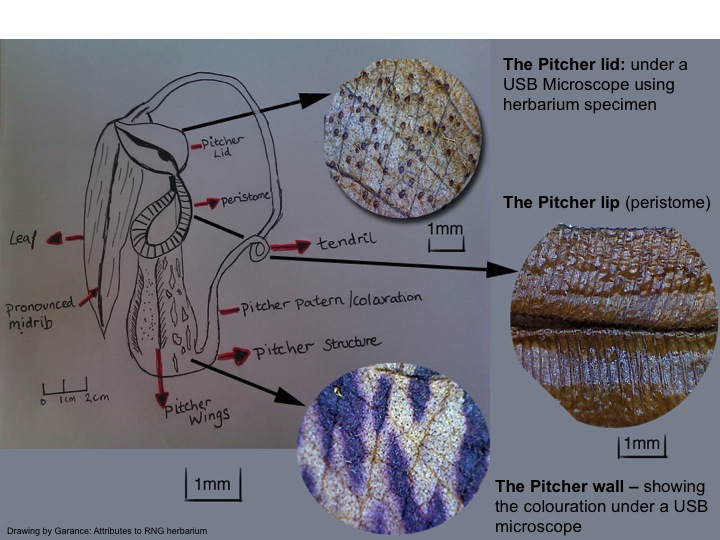
Picture 3: Drawing of a typical Pitcher structure with close ups using a USB microscope from herbarium specimens (RDG)
So what of the Taxonomy? First Stop: Generic Level.
Carnivorous plants have long been a source of intrigue and fascination right from the description made by Charles Darwin in his book Insectivorous Plants (Darwin 1875). Linnaeus named the genus Nepenthes in 1737 with far fewer species then there are now. The fascination with this genus which has shown incredible infraspecific variation making the task of delimitation harder for taxonomists especially with regards to the pitchers. It’s no wonder Nepenthaceae over the years has been a source of taxonomic confusion. The monographic revisions of Hooker (1873), MacFarlane (1908) and later the Dutch botanist Danser (1928) has not fully resolved the family. Danser was one of the first to propose a natural division of the genera Nepenthes into 6 groups based on morphological characters; Vulgatae, Montanae, Nobiles, Regiae, Insignes & Ureolatae. In total he recognised 65 species with circa 17 sp newly described taxa. Danser’s work greatly contributed towards other monographic creating a starting point to classify the Nepenthes species.
Second Stop: Order Level.
Nepenthes has not just been problematic on the infrageneric level but also on the order level too. Currently after advanced molecular and morphological analysis APG 3 has placed Nepenthaceae within the Caryophyllales (Non-Core) Clade (Stevens, 2007 and later). However, this has not always been the case. A previous classification by Cronquist (1988) has associated Nepentheceae with Droseraceae and Saraceniaceae in the order Nepenthales. This was on the basis that these families thrive in a low nutrient environment, while also sharing the carnivorous trait. Cronquist placed the order in the Subclass Dilleniidae. The Angiosperm Phylogeny group classification revealed its close links to Droseraceae, Diconcophyllaceae and Ancistrocladaceae, forming a clade within the order Caryophyllales. It has been suggested that the genus Triphyophyllum, placed in the Diconcophyllaceae family found growing in Africa is the link to this carnivorous lineage. Chase (1992) observed a similar convergence of morphological characters from both families being represented in this genus. This monotypic genus produces long lianas often up to 50m in length, similar to that of Nepenthes (Albert & Chase, 1992). In addition the midribs typically extends in a hook shape very much like the modified midrib forming pitchers in Nepenthes.
Interestingly, Darwin recognised that the carnivorous plants share different lineages which is very much in agreement to current day taxonomy. However, their interrelations and placement with other angiosperms has caused many contradictions. Darwin stated in his insectivorous plants book that Nepenthaceae were “not at all related to the Droseraceae” (Insectivorous plants, p. 292). However, we now know this is not the case, some Droseraceae species have similar rosette phase to that of Nepenthes and share the same vascularised glands with Triphyophyllum and Drosophyllum. Chase et al. (1992) looked at carnivorous plant evolution using phylogenetic analysis, and according to the research showed the carnivorous plants within the Caryophyllales to be monophyletic.
Although the Caryophyllales order contains a great proportion of the carnivorous lineages, the Sarraceniaceae that was previously thought to be closely linked to Nepenthes is now recognised to be part of the Ericales. The history of the Caryophyllales has not been straightforward either, and much more work is still being carried out to fully determine all of the interfamilial relationships and their placements. However, in-depth chloroplast sequencing (matK gene) did show there to be two distinct clades (Meimberg et al. 1999). One otherwise known as the Core Caryophyllales with main families; Polygonaceae, Plumbaginaceae, Tamaricaceae and Frankeniaceae, Simmondsiaceae. While the other group (non-core) contains all the carnivorous families; Nepenthaceae, Droseraceae and the sister taxa Drosophyllaceae, Dioncophyllaceae and Ancistrocladaceae.
This confirms the carnivorous genera are each monophyletic, especially Nepenthes which is the most ancient (Meimberg et al. 1999). It is quite remarkable how this family has successfully evolved and adapted, to be placed in an order which contains families that do not have carnivorous traits. This not only adds to their mysterious yet wonderful nature and will undoubtedly continue to intrigue Scientist and Botanists.
Key References
Pingback: Nepenthes mira – The Wonderful Pitcher plant | Tropical Biodiversity